LỘ TRÌNH “THẤU HIỂU SHIKIMATE – HÀNH TRÌNH ĐI TỪ ĐƯỜNG ĐẾN HƯƠNG”
Cây trồng không có bệnh, cây biết tự bảo vệ, tự điều chỉnh, tự làm đẹp, khi con đường này thông suốt. Con đường Shikimate — nếu hiểu thấu, chúng ta không chỉ nắm một “con đường sinh hóa”, mà là một bản đồ của sự sống, nơi đường → năng lượng → ý chí sinh tồn → hương vị → miễn dịch → sinh thái gắn chặt vào nhau của thực vật.
Để đi sâu – rộng – mở tư duy mọi ngóc ngách của nó, chúng ta cần thay đổi cách nhìn, cách hỏi, và cách kết nối. Cùng Seiko tìm hiểu bạn nhé.
I. Năng lượng đầu vào: từ ánh sáng đến “đường có ý nghĩa”
Trong Shikimate, đường (carbohydrate) không chỉ là nguồn năng lượng, mà là “nguyên liệu tri thức” để cây tự tổ chức lại cấu trúc sống.
- Nguyên liệu: Cây xanh sử dụng carbon dioxide (CO2) từ không khí và nước (H2O) từ đất.
- Cơ chế: Với sự xúc tác của ánh sáng mặt trời và chất diệp lục (chlorophyll) có trong lá, cây chuyển hóa các nguyên liệu này thành đường và giải phóng oxy (O2) vào khí quyển.
- Phương trình tổng quát: 6CO2+6H2O+Ánh sáng→C6H12O6 (glucose)+6O2
Từ glucose, cây tạo ra hai “đồng tiền” để mở Shikimate:
-
PEP (phosphoenolpyruvate) từ đường phân (glycolysis).
-
E4P (erythrose-4-phosphate) từ pentose phosphate pathway (PPP).
PEP + E4P gặp nhau (phản ứng DAHP synthase) → DAHP → Shikimate → Chorismate.
Từ chorismate, cây sinh ra ba amino thơm: Phe, Tyr, Trp (trung tâm của sắc – hương – vị – miễn dịch). -
👉 Khi cây tổng hợp PEP và E4P, đó là lúc ánh sáng (quang hợp) được chuyển hóa thành ý nghĩa sinh học.
1. Glucose tạo ra PEP (Phosphoenolpyruvate)
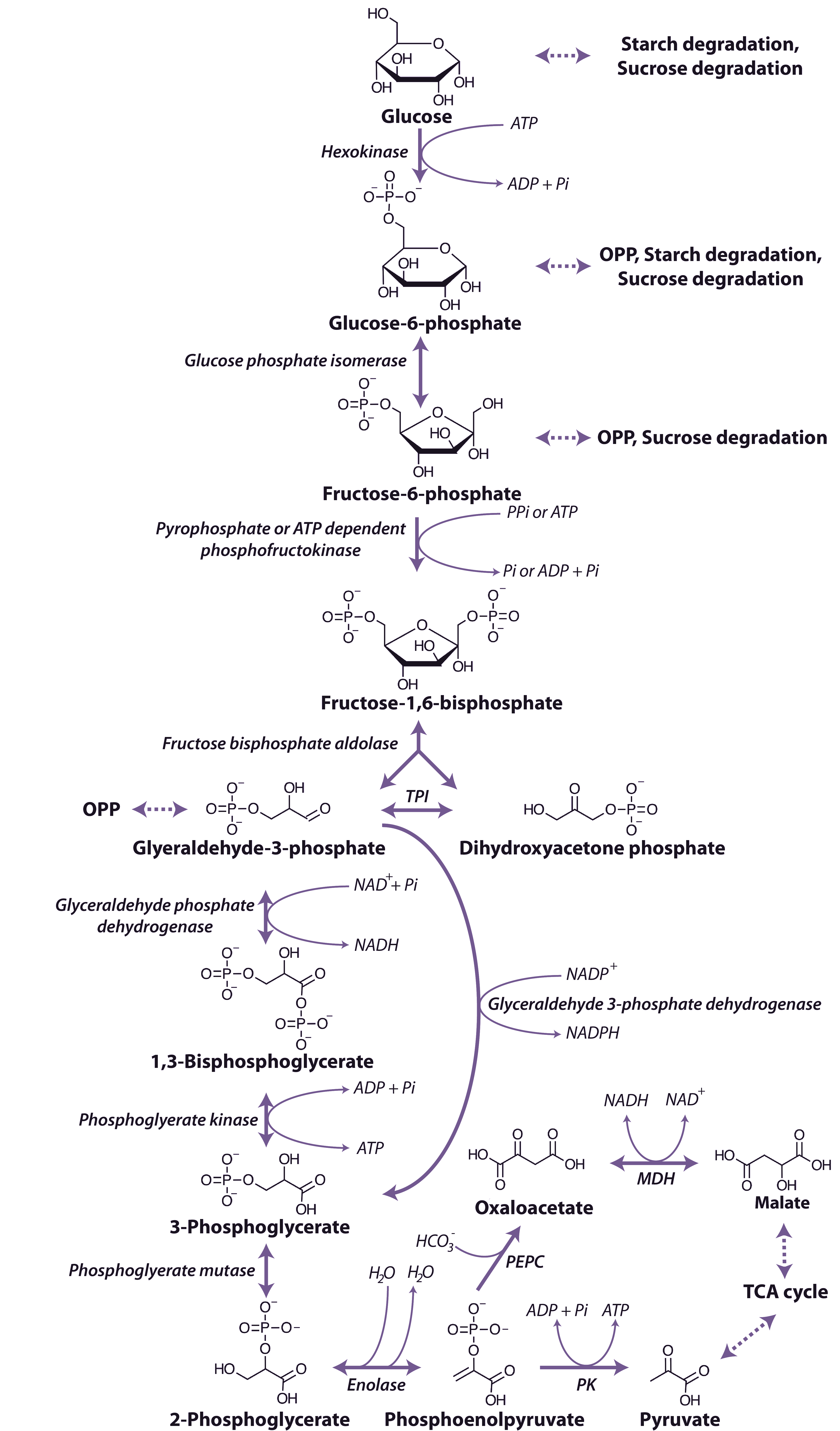
- Khởi đầu: Glucose đi vào con đường đường phân và được chuyển hóa qua nhiều bước trung gian.
- Các bước chính: Glucose được phosphoryl hóa thành Glucose-6-phosphate, rồi Fructose-6-phosphate, và sau đó phân cắt thành các phân tử 3 carbon.
- Tạo PEP: Sau nhiều phản ứng enzyme, một phân tử trung gian là 3-phosphoglycerate được biến đổi thành 2-phosphoglycerate, và cuối cùng, enzyme enolase loại bỏ một phân tử nước để tạo thành Phosphoenolpyruvate (PEP).
2. Glucose tạo ra E4P (Erythrose-4-phosphate)
E4P được tạo ra chủ yếu thông qua Chu trình Pentose Phosphate (PPP):
- Con đường Oxidative (Oxy hóa): Glucose-6-phosphate (sản phẩm đầu tiên của đường phân) được tách ra khỏi con đường chính để đi vào chu trình PPP.
- Tạo E4P: Trong nhánh không oxy hóa của chu trình PPP, một loạt các phản ứng tái sắp xếp carbon xảy ra, trong đó enzyme transaldolase và transketolase tham gia biến đổi các đường trung gian (như Fructose-6-phosphate và Glyceraldehyde-3-phosphate) để tạo ra Erythrose-4-phosphate (E4P).
–
3. Vì sao cây cần tổng hợp PEP và E4P?
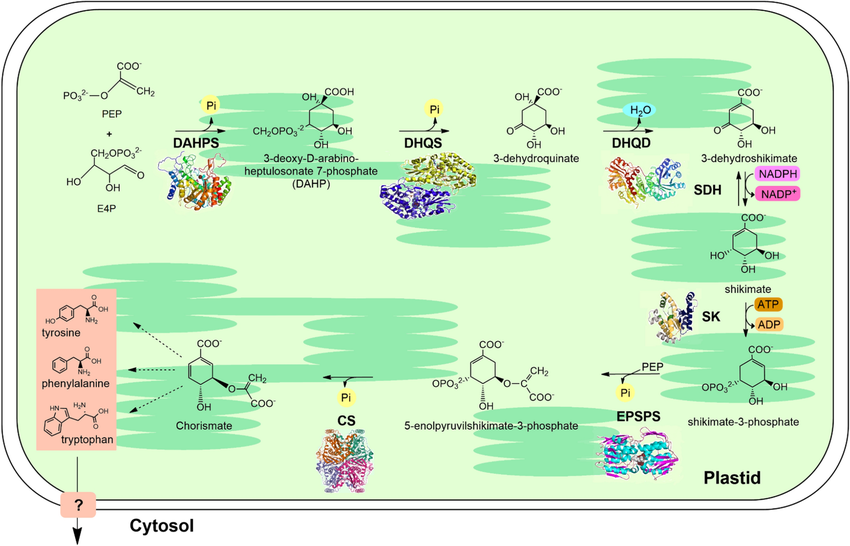
- Phenylalanine (Phe)→ Flavonoid, Lignin, SA (kháng bệnh, cứng mô, màu đỏ tím)
- Tyrosine (Tyr) → Alkaloid, chống oxy hóa, dẫn truyền tín hiệu stress
- Tryptophan (Trp)→ Auxin (rễ, chồi, quả), Melatonin, Indole alkaloid (phòng vệ, hương vị)
Đây chính là 3 hướng của sự sống:
-
Một hướng bảo vệ (defense)
-
Một hướng phát triển (growth)
-
Một hướng biểu hiện (expression: màu, mùi, vị)
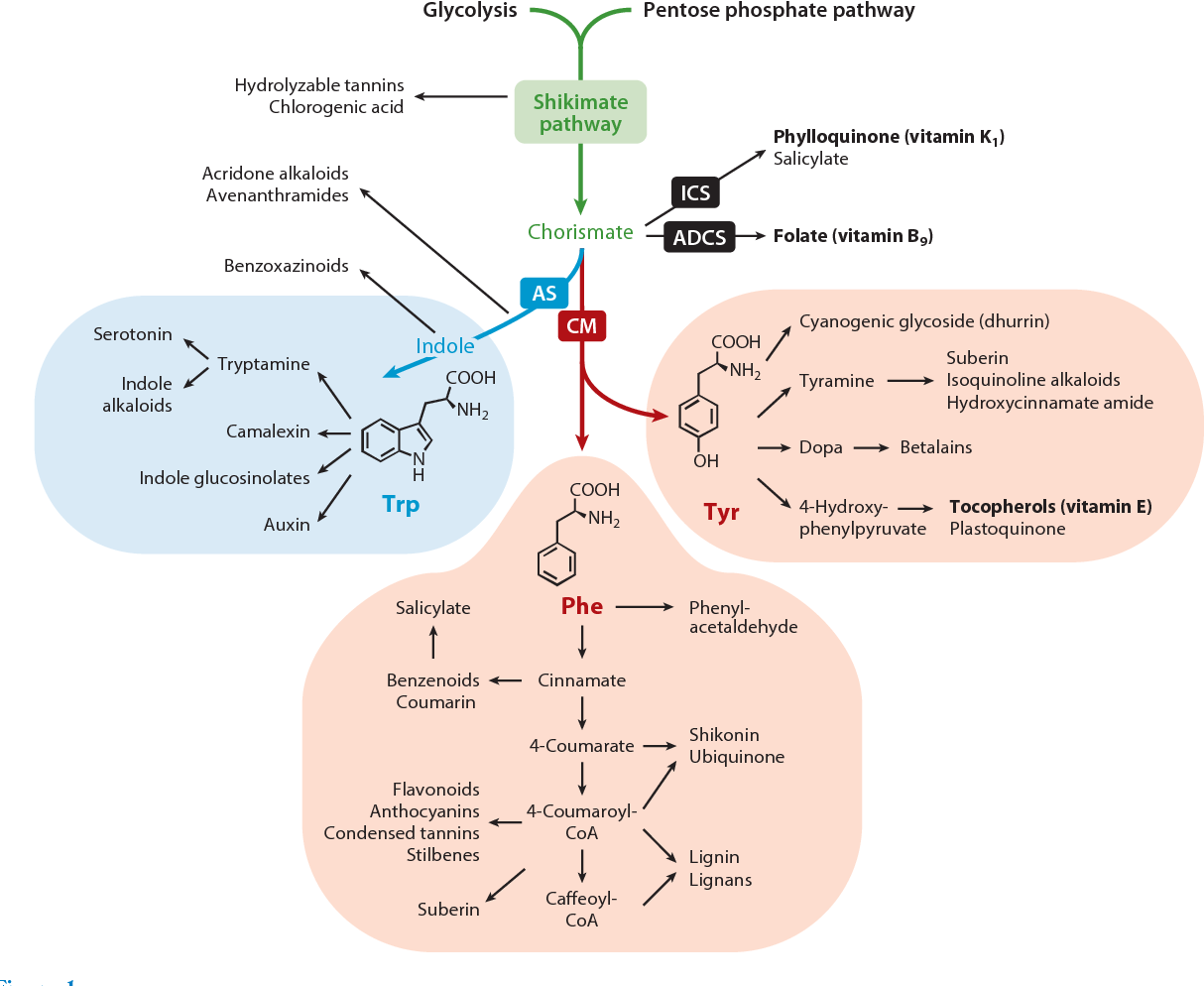
-
Tóm lại, PEP và E4P là những mắt xích trung tâm kết nối quá trình chuyển hóa năng lượng chính (đường phân, quang hợp) với quá trình sản xuất các hợp chất bảo vệ và cấu trúc thiết yếu cho sự tồn tại và phát triển của cây trồng.
Cách tư duy ở đây là:
“Đường không chỉ nuôi cây, mà giúp cây biết mình là ai.”
➡️ Nếu cây đói năng lượng (thiếu ánh sáng, thiếu Mg, thiếu P) → Shikimate yếu → cây mất trí nhớ sinh học → không tạo được sắc, hương, vị, và miễn dịch.
–
II. Ba nhánh Shikimate: “ba dòng sinh mệnh”
-
NHÁNH PHENYLALANINE (Phe) – “BỘ XƯƠNG & HỆ MIỄN DỊCH” CỦA CÂY
1.1. Enzyme then chốt:
-
PAL (Phenylalanine Ammonia Lyase): tách nhóm NH₂ khỏi Phe để tạo cinnamic acid, là “cửa ngõ vàng” mở ra toàn bộ ngành công nghiệp phenolic của cây.
-
Từ cinnamate → 4-coumarate → 4-coumaroyl-CoA → nhiều nhánh sản phẩm.
1.2. Chuỗi sinh hóa và sản phẩm:
| Gốc từ Phe | Sản phẩm tiếp theo | Vai trò |
|---|---|---|
| Cinnamic acid → 4-Coumarate → Coumaroyl-CoA | Flavonoid, Anthocyanin, Stilbene | Màu sắc (đỏ, tím, xanh), chống tia UV, bảo vệ DNA, tăng hương vị |
| Caffeoyl-CoA | Lignin, Lignan, Suberin | Làm cứng mô, chống đổ ngã, chống xâm nhập nấm – vi khuẩn, khoá cửa nấm, côn trùng |
| Benzeneoid, Coumarin, Phenylacetaldehyde | Hương thơm hoa quả (hoa nhài, cam, tiêu, cà phê) | Thu hút thụ phấn, tăng chất lượng nông sản |
| Salicylic acid (SA) | Hormone miễn dịch thực vật | Kích hoạt SAR – “vaccine nội sinh” chống bệnh |
1.3. Kết nối Shikimate ↔ Miễn dịch:
-
Khi vi sinh hoặc nấm tấn công, cây kích hoạt PAL → tăng dòng Phe → SA tăng → SAR (Systemic Acquired Resistance).
-
SA thúc đẩy biểu hiện gene PR (pathogenesis-related proteins) → enzyme kháng bệnh (POD, PPO, β-1,3-glucanase…).
-
Khi Phe↔PAL mạnh, SA nội sinh cao → gene PR bật → vách tế bào cứng, men kháng bệnh tăng → ít cần thuốc.
1.4. Ứng dụng nông học:
-
Giữ Mg, Fe, P đầy đủ → Shikimate mạnh → PAL hoạt động tốt.
-
Bổ sung amino acid Phe hoặc humic–fulvic → tăng tổng hợp phenolic.
-
Khi lá có ánh tím nhẹ, màu quả đậm, mô cứng, nghĩa là Phe đang “hát”.
–
2. TYROSINE (Tyr) — “BỘ NÃO & HỆ DẪN TRUYỀN NĂNG LƯỢNG”
2.1. Enzyme then chốt:
-
Tyrosine aminotransferase, 4-hydroxyphenylpyruvate dioxygenase (HPPD), và tyramine decarboxylase (TDC) – là 3 điểm chuyển hóa trung tâm.
-
Tạo ra các hợp chất phenolic và alkaloid giàu năng lượng redox.
2.2. Chuỗi sinh hóa & vai trò:
| Gốc Tyr | Sản phẩm | |
|---|---|---|
| 4-Hydroxyphenylpyruvate → Tocopherol (Vitamin E) | Chống oxy hóa màng, bảo vệ diệp lục, bền màu lá | |
| DOPA → Betalain (màu đỏ, tím ở củ dền, xương rồng) | Màu sắc tự nhiên, bảo vệ khỏi tia UV | |
| Tyramine, Isoquinoline alkaloid | Kháng nấm, kháng khuẩn, truyền tín hiệu stress, kháng nấm khuẩn, làm kín vỏ–rễ. | |
| Hydroxycinnamate amide, Suberin | Tăng cứng mô, làm màng bảo vệ rễ – thân – quả | |
| Plastoquinone (từ Tocopherol nhánh) | Tham gia chuỗi quang hợp, vận chuyển electron |
2.3. Tư duy sinh học:
Tyr là nguồn của tín hiệu chống stress.
Khi cây bị nóng, thiếu nước, độc nhôm hoặc thuốc, hoạt tính HPPD và TDC tăng → tạo catecholamine thực vật (dopamine, tyramine) → cây “bình tĩnh” và phục hồi nhanh sau sốc nhiệt/khô hạn/thuốc..
2.4. Ứng dụng thực tế:
-
Tăng Tyr bằng amino acid hoặc bổ sung Fe, Mn, Zn để enzyme HPPD hoạt động.
-
Trong stress nóng hoặc thiếu nước → phun HaifaStim Energy giúp cây sinh catecholamine tự nhiên → hạn chế sốc.
-
Khi lá bền, xanh, ít cháy nắng → Tyr đang hoạt động mạnh.
–
3. TRYPTOPHAN (Trp) — “TIM, HORMONE & HƯƠNG VỊ CỦA CÂY”
3.1. Enzyme then chốt:
-
Tryptophan aminotransferase (TAA1),
-
Indole-3-pyruvate decarboxylase (YUC family) – hai enzyme chính chuyển Trp thành IAA (auxin).
-
Các nhánh khác dẫn đến indole alkaloid, serotonin, melatonin.
3.2. Chuỗi sinh hóa & vai trò:
| Gốc Trp | Sản phẩm | Vai trò |
|---|---|---|
| Indole-3-acetic acid (IAA) | Hormone tăng trưởng – kích rễ, chồi, quả | Kiểm soát cân bằng sinh trưởng |
| Indole alkaloid (camalexin, benzoxazinoid) | Chống nấm, vi khuẩn, tuyến trùng | Phòng vệ sinh học |
| Serotonin, Tryptamine | Ổn định nhịp sinh học, điều hòa stress | Hỗ trợ quang hợp và đậu trái |
| Melatonin thực vật | Đồng hồ sinh học, điều tiết đêm–ngày | Cải thiện năng lượng và sức đề kháng |
3.3. Ý nghĩa sinh lý học:
Trp là “nguồn hormone và cảm xúc của cây”.
Khi ánh sáng – nước – đạm cân bằng → Trp chuyển hóa thuận → rễ trắng, chồi vươn, hoa đồng đều.
Khi stress hoặc hóa chất phá vỡ Shikimate → Trp nghẽn → cây “câm” (rễ không ra, hoa không nở).
3.4. Ứng dụng thực hành:
-
Giai đoạn ra rễ, ra hoa, đậu trái → phun HaifaStim Energy (Trp + amino tự do cao).
-
Kết hợp Ca + B + Zn + K để Auxin vận hành ổn định.
-
Dưới mưa kéo dài, thiếu sáng → duy trì Trp giúp giữ cân bằng hormone – tránh rụng hoa.
Bạn đọc thêm để hiểu hơn về Auxin ở đây nhé: https://seikoagri.com.vn/co-che-tong-hop-auxin-iaa/
Phe = miễn dịch & cấu trúc
Tyr = nhận thức & phản xạ
Trp = hormone & sinh sản
Ba nhánh gặp nhau ở Chorismate, nơi năng lượng quang hợp và carbon “nảy sinh ý thức sinh học” của cây.
Cây trồng không có bệnh, cây biết tự bảo vệ, tự điều chỉnh, tự làm đẹp, khi con đường này thông suốt.
–
III. Kết nối ngược: vì sao Shikimate “chết lâm sàng” khi đất – lá – đạm lệch?
3.1 Shikimate – Nút giao giữa năng lượng và sinh học
Vị trí: Shikimate pathway nằm ở lục lạp (chloroplast) – nơi vừa diễn ra quang hợp, vừa liên thông với chu trình đường phân (glycolysis) và pentose phosphate pathway (PPP).
Hai nguyên liệu khởi đầu:
-
Phosphoenolpyruvate (PEP) – từ đường phân (glycolysis) trong tế bào chất.
-
Erythrose-4-phosphate (E4P) – từ Pentose phosphate pathway.
Để hai phân tử này gặp nhau và bắt đầu chu trình Shikimate, cây cần 3 điều kiện sống:
👉 Có O₂ để hô hấp rễ
👉 Có ánh sáng để quang hợp
👉 Có lân hoạt hóa (Pi) để tái tạo ATP và NADPH.
Khi một trong ba yếu tố này bị nghẽn, Shikimate sẽ “chết lâm sàng”.
Khi cây bị stress, ví dụ thiếu oxy trong đất, rễ ngừng hô hấp → NADPH mất cân bằng → PEP không được tái tạo → Shikimate chậm lại.
Do đó, muốn bật lại Shikimate, phải:
-
Làm đất thoáng khí (tắt kỵ khí)
-
Duy trì quang hợp (Mg, Fe, Mn đủ)
-
Giữ lân ở dạng hoạt hóa (MKP, MAP, humic hỗ trợ)
👉 Shikimate là bản giao hưởng giữa khí – lân – ánh sáng – carbon.
–
3.2. HỆ HÔ HẤP RỄ – “NHỊP THỞ DƯỚI ĐẤT” (đất có O₂) → PEP mới chảy
1. Khi thiếu oxy trong đất:
-
Rễ ngừng hô hấp hiếu khí, không tạo được ATP và NADPH.
-
Quá trình đường phân bị kẹt ở pyruvate, không chuyển thành PEP được.
-
Vi sinh vật yếm khí chiếm ưu thế → sinh H₂S, CH₄, NH₃ → phá hủy ty thể rễ.
Kết quả:
→ PEP giảm, Shikimate chậm lại,
→ Không tổng hợp được phenylalanine → cây mềm mô, dễ bệnh, lá mỏng, thiếu sắc tố.
2. Dấu hiệu thực tế:
-
Đất có mùi “trứng thối” → H₂S → Shikimate tắc
- Rễ nâu đen, đất “bết – dẽ
-
Đất thơm nhẹ, tơi, rễ trắng → Hô hấp mở → Shikimate hoạt động.
3. Giải pháp:
| Mục tiêu | Biện pháp |
|---|---|
| Khôi phục hô hấp rễ | Xới phá váng, thoát nước, phủ rơm thoáng khí |
| Kích hoạt vi sinh hiếu khí | Tưới EM, humic, Ca(NO₃)₂ nồng độ thấp (NO₃ giúp rễ hô hấp “thay O₂”) |
| Tăng “lưu thông khí đất” | Phủ Hữu cơ Seiko Organic hoặc hữu cơ có cấu trúc sợi (không ủ tươi), không nén chặt gốc |
–
III. HỆ QUANG HỢP – “BỘ MÁY TẠO NĂNG LƯỢNG VÀ CARBON”
Quang hợp (lá đủ Mg–Fe–Mn & ánh sáng) → NADPH/ATP đủ để khử
1. Năng lượng Shikimate cần:
-
ATP để cung cấp phosphate cho các phản ứng phosphoryl hóa.
-
NADPH để khử 3-dehydroshikimate → shikimate → chorismate.
Cả ATP và NADPH đến từ pha sáng quang hợp,
nên khi ánh sáng yếu, hoặc thiếu Mg, Fe, Mn, cây mất năng lượng “khử”,
→ Shikimate không thể đi đến sản phẩm cuối (Phe, Tyr, Trp).
2. Tầm quan trọng của nguyên tố:
| Nguyên tố | Vai trò trong quang hợp | Hậu quả khi thiếu |
|---|---|---|
| Mg | Trung tâm của chlorophyll, vận hành Rubisco | Lá vàng nhạt, mất cân bằng NADPH |
| Fe | Thành phần ferredoxin, chuyển electron | Tắc chuỗi quang hợp, giảm năng lượng khử |
| Mn | Kích hoạt enzyme tách nước, khởi động quang hợp | Quá trình oxy hóa nước yếu, thiếu O₂ nội sinh |
➡️ Khi thiếu các nguyên tố này, chuỗi chuyền electron bị gián đoạn, Shikimate không được “tiếp điện”.
3. Giải pháp:
-
Giữ lá xanh – dày – sáng bằng Mg(NO₃)₂, Fe-EDDHA, Mn chelate.
-
Phun HaifaStim Force (mannitol + betaine) giúp ổn định diệp lục trong nắng nóng, tránh “tắc NADPH”.
-
Duy trì ánh sáng, tránh che phủ quá dày.
–
IV. HỆ NITRAT – “NHỊP SINH HỌC CỦA ĐẠM VÀ CARBON”
Nhịp N (NO₃⁻/NH₄⁺/Amino) → C:N cân
1. Vai trò cân bằng C:N trong Shikimate:
Shikimate pathway phụ thuộc carbon cố định (C), trong khi đạm (N) đi vào qua các amino acid (Phe, Tyr, Trp).
Nếu cây thừa đạm – thiếu carbon, quá trình bị “lệch nhịp”:
-
PEP và E4P bị hút sang chu trình ammonium assimilation.
-
Shikimate yếu đi → cây mềm mô, xanh nước, ít phenolic → dễ bệnh.
2. Dạng đạm ảnh hưởng:
| Dạng đạm | Tác động đến Shikimate |
|---|---|
| NH₄⁺ (amoni) | Giảm pH rễ, giảm oxy, ức chế hô hấp → Shikimate giảm |
| NO₃⁻ (nitrat) | Thúc đẩy hô hấp, tăng hoạt enzyme NADPH–nitrate reductase → Shikimate mạnh |
| Amino (hữu cơ) | Cung cấp N + carbon cùng lúc → giúp cây duy trì cân bằng C:N |
3. Giải pháp:
-
Ưu tiên đạm NO₃⁻ (Ca(NO₃)₂, Mg(NO₃)₂) thay vì NH₄⁺.
-
Bổ sung amino acid tự nhiên (HaifaStim Energy) để đồng bộ hóa N và C.
-
Giữ đất “có khí” để vi sinh nitrat hóa hoạt động (tạo NO₃⁻ đều đặn).
–
V. Lân hoạt hóa (Pi) → khóa ATP/NADPH, mở DAHP
-
P bị khóa (al–Fe–Ca phosphate) ⇒ DAHP synthase thiếu Pi ⇒ nghẽn ngay cửa vào.
-
Sửa: MKP/MAP liều hợp lý + humic/fulvic (giữ Pi hoạt hóa vùng rễ).
Kết luận phần nối hệ: Shikimate = giao hưởng giữa khí (O₂) – lân (Pi) – ánh sáng – carbon.
Một “nốt” lệch, cả bản nhạc tắt.
| Hệ thống | Khi hoạt động tốt | Khi bị nghẽn |
|---|---|---|
| Hô hấp đất | PEP tăng → Shikimate khởi động | Thiếu O₂, rễ thâm đen, H₂S cao |
| Quang hợp | NADPH + ATP dồi dào → chorismate tạo mạnh | Thiếu Mg/Fe/Mn, lá úa, diệp lục yếu |
| Nitrat – C:N | Carbon & Nitrogen đồng nhịp → phenolic, flavonoid tăng | Thừa NH₄⁺, thiếu C, mất màu quả |
| Lân hoạt hóa | MKP, MAP, humic giúp giữ Pi ổn định trong rễ | Đất chua, P bị khóa → Shikimate “đứt mạch” |
🌱 Khi đất thở – lá quang hợp mạnh – nitrat ổn định – lân hoạt hóa,
Shikimate sẽ “hát lên”, cây bước sang trạng thái thông minh sinh học.
–
VI. THỰC HÀNH : “KHỞI ĐỘNG LẠI SHIKIMATE TRONG 7 NGÀY”
-
Ngày 1–2: Xới phá váng, tưới Ca(NO₃)₂ 0,3% để mở O₂ và nitrat hóa đất.
-
Ngày 3–4: Phun Mg(NO₃)₂ + HaifaStim Force để kích quang hợp, nạp năng lượng khử.
-
Ngày 5: Bổ sung MKP (0-52-34) liều nhẹ + humic → giữ lân hoạt hóa cho PEP–E4P.
-
Ngày 6–7: Phun HaifaStim Energy (Amino + Trp) để nối Shikimate → auxin–flavonoid.
→ Sau 7 ngày: lá xanh sâu, rễ trắng mạnh, lá dày bóng — tín hiệu Shikimate hoạt hóa lại.
Bảng kiểm tra “làm – biết – sửa” (cheat-sheet ngoài đồng)
| Hiện tượng | Nghẽn ở đâu | Việc cần làm |
|---|---|---|
| Đất hôi, rễ nâu | Hô hấp rễ (thiếu O₂) | Thoát nước, xới nhẹ đất, phá váng, NO₃⁻ loãng, EM + humic |
| Lá xanh nước, dễ bệnh | Shikimate thiếu phenolic (Phe thấp), thừa NH₄⁺ | Giảm NH₄⁺, tăng NO₃⁻, bổ sung MKP + Mg/Fe/Mn, biostimulant Haifastim Energy |
| Lá úa, dễ cháy nắng | Thiếu Mg/Fe/Mn → thiếu NADPH/ATP | Phun Mg(NO₃)₂ + Fe-EDDHA + vi lượng có Mn; giữ tán sáng |
| Rễ chậm, hoa dễ rụng | Trp → IAA nghẽn | Amino tự do (Trp), Ca+B+Zn+K đúng nhịp |
-
Nguồn lân hoạt hóa: MKP (0-52-34), MAP (12-61-0) liều vừa, đều.
-
Nguồn nitrat sạch: Ca(NO₃)₂, Mg(NO₃)₂.
-
Vi lượng quang hợp: Mg, Fe-EDDHA, Mn.
-
Chất điều hòa–bảo vệ: HaifaStim Force (amino + mannitol + betaine), HaifaStim Energy (amino tự do/Trp), Seiko Humic Mỹ (humic/fulvic).
Chúng ta cần ghi nhớ ngắn gọn như sau:
-
“Đất thở trước – lá sáng sau – đạm theo nhịp – lân luôn mở.”
-
“PEP + E4P là cửa; Phe–Tyr–Trp là ba nhánh sống.”
-
“Phe dựng khiên – Tyr giữ thần – Trp nắn hormone.”
VII. Hỏi- Đáp
-
Nếu Shikimate dừng lại, phản ứng nào trong đất sẽ bù vào?
Câu hỏi này chúng ta sẽ làm rõ “Liên hệ vi sinh và mối quan hệ đất–rễ”.
Khi nó dừng → dòng carbon từ PEP & E4P không đi vào “hệ thống tri thức” nữa, mà bị “đổ sang” các phản ứng thay thế, giúp cây duy trì năng lượng tạm thời, nhưng mất chức năng sinh học cao cấp (miễn dịch, màu, hương, lignin).
Trong đất và rễ: 5 con đường “bù” cho Shikimate (các phản ứng thay thế)
(1) Fermentation (lên men) – phản ứng “thở giả” trong điều kiện kỵ khí
-
Khi đất thiếu oxy, rễ không hô hấp hiếu khí, pyruvate không vào TCA (Krebs) mà chuyển sang lactic acid hoặc ethanol fermentation.
-
Kết quả:
-
ATP vẫn được sinh nhưng rất ít (2 ATP thay vì 38 ATP).
-
Môi trường acid hóa, tích tụ ethanol, acid hữu cơ, rễ bị stress.
-
Vi sinh yếm khí (Clostridium, Desulfovibrio…) tăng → sinh H₂S, CH₄, NH₃.
-
Hiểu theo sinh thái:
Đất bắt đầu “thở bằng miệng”, không bằng phổi → có năng lượng, nhưng không có ý thức sinh học → Shikimate tắt, rễ đen, cây vẫn xanh nhưng “chết cảm xúc”.
–
(2) Methanogenesis & Sulfate reduction – “bù năng lượng” bằng phản ứng khử
-
Khi O₂ mất, vi sinh yếm khí lấy electron từ carbon hữu cơ để khử:
-
CO₂ → CH₄ (methane)
-
SO₄²⁻ → H₂S (hydrogen sulfide)
-
-
Hai khí này sinh ra năng lượng cho vi sinh, chứ không cho cây.
-
Nhưng cây lại chịu độc:
H₂S ức chế enzyme cytochrome oxidase trong ty thể rễ → rễ ngừng hô hấp, ATP giảm → PEP không tái tạo → Shikimate không khởi động lại được.
🌋 Đây là lý do đất “bốc mùi trứng thối”, lá cây xanh nhưng mỏng, mau héo, bệnh rễ lan.
–
(3) Nitrate reduction → Ammonium assimilation – “cân đạm” bù năng lượng
-
Khi cây thiếu carbon (do Shikimate tắt), nó chuyển hướng hấp thu NH₄⁺ nhiều hơn, vì NH₄⁺ dễ đồng hóa, ít tốn ATP hơn NO₃⁻.
-
Nhưng NH₄⁺ đi vào rễ quá nhiều →:
-
pH vùng rễ giảm,
-
ty thể giảm hoạt tính,
-
vi sinh nitrat hóa chết.
-
Kết quả: rễ hô hấp kém, N bị “kẹt”, carbon không đủ → lá xanh đậm, thân mềm, bệnh nhiều.
⚠️ Đây chính là kiểu “xanh hóa học” mà nông dân tưởng là khỏe, nhưng thật ra Shikimate đã chết lâm sàng.
–
(4) Polyamine synthesis – cây “câu giờ” bằng hormone stress
-
Khi Tyr và Trp không được tổng hợp → cây tăng tổng hợp putrescine, spermidine, spermine (polyamine).
-
Chúng là hormone stress, giúp tạm giữ ổn định màng, chống oxy hóa.
-
Tuy nhiên: nếu Shikimate không khởi động lại → polyamine tích tụ → ức chế quang hợp, ngộ độc nội sinh.
Giống như cơ thể người dùng adrenaline để sống sót khi đói, nhưng không thể sống mãi trong stress.
–
(5) Lên men humic – vi sinh heterotrophic chiếm quyền
-
Khi cây không điều phối carbon qua Shikimate, lượng đường rỉ rễ (root exudate) thay đổi:
-
Vi sinh kỵ khí – phân hủy protein – sinh NH₃ tăng,
-
Vi sinh cố định carbon – nitrat hóa – sinh enzyme Shikimate-like giảm.
-
-
Đất trở thành “xưởng lên men” thay vì “phòng thí nghiệm Shikimate”.
Đó là lý do sau mưa, đất có mùi hôi, vi sinh tốt giảm, nấm bệnh tăng — Shikimate ở mức đất cũng “tắt”.
–
Cây phản ứng thế nào?
| Cấp phản ứng | Biểu hiện | Hệ quả |
|---|---|---|
| Tế bào | Mất NADPH/NADP⁺ cân bằng | Enzyme khử 3-dehydroshikimate ngừng |
| Rễ | Pyruvate không vào TCA | PEP giảm, rễ không trắng |
| Lá | Thiếu ATP/NADPH | Quang hợp giả (xanh không tạo chất) |
| Cây tổng thể | Giảm Phe, Tyr, Trp | Mất lignin, flavonoid, auxin, SA |
| Đồng ruộng | Lá mỏng, quả nhạt, bệnh lá lan | Mất “ý thức sinh học” |
–
Cách bật lại Shikimate sau khi nó dừng
Muốn cây sống thật, không chỉ xanh – ta phải khôi phục tuần hoàn năng lượng thật:
-
Tắt phản ứng methanogenesis & H₂S: phá váng, thoáng khí, Ca(NO₃)₂ 0,3%.
-
Bật hô hấp – ATP nội sinh: EM hiếu khí, humic, pH vùng rễ 6–6.5.
-
Bật quang hợp: Mg–Fe–Mn; lá phải sáng, dày, không “xanh nước”.
-
Mở lân hoạt hóa: MKP + humic (VIM/ Seiko Humic)
-
Cung cấp amino “gốc Trp–Phe–Tyr” → HaifaStim Energy + Force luân phiên 3–5 ngày.
-
Giữ nitrat ổn định: không để NH₄⁺ vượt NO₃⁻ quá cao.
Khi Shikimate sống lại, đất không còn mùi CH₄ – H₂S, rễ tái trắng, và cây chuyển từ “xanh giả” sang “xanh bền”.
Tư duy tổng hợp: Shikimate = đường trí tuệ
Khi Shikimate hoạt động:
-
Carbon → sắc màu, hương, trí nhớ miễn dịch.
-
Đất → “thở” bằng O₂.
-
Cây → “sống” bằng ý thức sinh học.
Khi Shikimate tắt:
-
Carbon → lên men, khí độc, năng lượng rẻ tiền.
-
Cây → “thở bằng miệng”, xanh giả, nhanh tàn.
🪶 Đó là ranh giới giữa nông nghiệp hóa học và nông nghiệp sinh học.
Cùng một cây, một lá – khác nhau ở việc Shikimate còn hoạt động hay không.
2. Nếu tôi kích hoạt Shikimate bằng nguồn carbon ngoài (như amino, mannitol, humic), cây có nhớ không?
Có – nhưng phải đúng cách.
Còn nếu sai — cây chỉ “sống vay mượn”, rồi quên mất cách sống của chính mình.
Giờ ta đi thật sâu, từng lớp một, để hiểu cơ chế “nhớ – quên – tự lập năng lượng” của Shikimate.
Cây có trí nhớ sinh học — và Shikimate là nền thần kinh trung ương của nó
Cây không có não, nhưng có mạng lưới trí nhớ phân tử:
-
ở ty thể (hô hấp) – ghi nhớ năng lượng,
-
ở lục lạp (quang hợp) – ghi nhớ ánh sáng,
-
ở nhân & màng tế bào – ghi nhớ tín hiệu miễn dịch.
→ Shikimate nằm ngay giao điểm của ba hệ đó:
Nó “nhớ” được khi:
-
Có chu kỳ năng lượng đầy đủ (ATP–NADPH)
-
Có nguồn carbon nội sinh (từ quang hợp)
-
Có enzyme “cảm nhận” tín hiệu môi trường hoạt động nhịp nhàng.
Khi ba điều này đủ, cây học cách tái tạo Phe–Tyr–Trp → nó không cần ta “nuôi” nữa, mà tự bật Shikimate.
–
2. Khi ta đưa carbon ngoài vào – cây “nhận viện trợ”
a. Amino acid (đặc biệt là Trp, Phe, Gly)
-
Là dạng carbon–nitrogen cân bằng, dễ hấp thu.
-
Khi hấp thu qua lá hoặc rễ, cây giảm tải đường phân (glycolysis) → đỡ tiêu năng lượng.
-
Nếu chỉ bổ sung ngắn hạn (1–3 ngày), amino giúp tái nạp NADPH và “gợi nhớ” cho enzyme Shikimate.
👉 Kết quả tốt: cây phục hồi, rễ bật, lá dày.
👉 Nhưng nếu dùng quá lâu: cây quen ăn sẵn, ức chế enzyme nội sinh → “mất trí nhớ năng lượng.”
Giống như người tập thể dục mà ngày nào cũng uống nước đường: khỏe tạm, nhưng cơ thể quên cách tạo ATP.
–
b. Mannitol, betaine – “đường thông minh”
-
Mannitol là polyol tự nhiên, cây tạo ra khi stress (thiếu nước, mặn, lạnh).
-
Nó ổn định NADPH/NADP⁺ ratio, tái hoạt hóa các enzyme khử trong Shikimate.
-
Không “làm thay” quang hợp, mà hỗ trợ tế bào tự điều hòa điện tử → giúp nhớ cơ chế “khử” năng lượng.
👉 Kết quả: cây hồi năng lượng mà không mất trí nhớ.
👉 Dùng đúng liều thấp, định kỳ 5–7 ngày là đào tạo chứ không thay thế.
–
c. Humic & fulvic – “thầy dạy hô hấp”
-
Humic không phải năng lượng, mà là bộ nhớ môi trường.
-
Nó kích hoạt enzyme dehydrogenase, catalase, polyphenol oxidase,
giúp rễ nhớ lại cách hô hấp hiếu khí và bật dòng PEP–E4P nội sinh. -
Còn fulvic (phân tử nhỏ hơn) gắn carbon hoạt hóa vào chloroplast, nối carbon quang hợp và carbon đất.
👉 Dùng humic đúng cách = gợi lại trí nhớ sinh học của đất.
👉 Quá liều = bão carbon → vi sinh yếm khí lợi dụng → rễ “lười thở”.
–
Vậy cây nhớ bằng gì?
Không phải bằng não, mà bằng:
-
Methylation & acetylation trong DNA – giống “ghi chú” cho enzyme hoạt động.
-
Histone code & redox memory – các protein “bật/tắt” gen Shikimate khi nhận tín hiệu NADPH, SA, ROS.
-
MiRNA – truyền “thói quen năng lượng” từ lá → rễ → thế hệ sau (thật, có chứng minh: SA và auxin có thể truyền ký ức phản ứng stress sang đời F1).
👉 Khi Shikimate được kích hoạt tự nhiên (qua ánh sáng, đất thở, Mg–Fe–P đầy đủ),
cây tự ghi nhớ → vụ sau gặp stress, phản ứng nhanh và nhẹ hơn.Nhưng nếu Shikimate chỉ được “ép chạy” bằng amino–đường ngoài, cây phục hồi nhanh mà quên nhanh.
–
Cách dùng nguồn carbon ngoài để “gợi nhớ”, không “ép nhớ”
| Mục tiêu | Biện pháp |
|---|---|
| Kích hoạt tạm thời | 1–2 lần amino + mannitol khi đất thoáng, sáng tốt |
| Gợi nhớ dài hạn | Duy trì humic + Mg–Fe–Mn + nitrat ổn định |
| Huấn luyện phản xạ | Giảm dần hỗ trợ ngoài, cho cây “tự đói – tự ăn” trong 5–7 ngày |
| Duy trì trí nhớ | Xen kẽ ẩm–khô – sáng–tối để enzyme thích nghi tự nhiên |
–
Sinh tồn – bài học của sự sống trong đất
Kích hoạt Shikimate đúng cách là dạy cây nhớ lại “ý chí sinh tồn” của nó.
Amino, mannitol, humic chỉ là chìa khóa, không phải “động cơ”.
Mục tiêu không phải cho cây khỏe tức thời, mà đánh thức bản năng thông minh sinh học.
Cây có nhớ.
Nhưng trí nhớ đó chỉ tồn tại khi ta cho nó cơ hội “tự bật đèn” —
không quá sáng, không quá tối, mà vừa đủ để tự chiếu sáng chính mình.
3. Khi cây ra hoa, Shikimate ưu tiên nhánh nào? Phe hay Trp?
Để trả lời câu hỏi này, chúng ta cùng làm rõ sự chuyển dịch năng lượng sinh sản nhé:
Nguyên tắc nền: Shikimate không “chọn bừa”
Cây tự điều phối dòng carbon–nitrogen theo mục tiêu sinh học ở từng giai đoạn:
-
Sinh trưởng (growth): hướng sang Tyr → năng lượng – quang hợp.
-
Bảo vệ (defense): hướng sang Phe → cấu trúc – kháng bệnh.
-
Sinh sản (reproduction): hướng sang Trp → hormone – tín hiệu hoa – quả.
Vì vậy, khi cây bước vào pha ra hoa, Shikimate chuyển dần “trọng lực” từ Phe sang Trp,
nhưng không tắt Phe hoàn toàn – vì hoa vẫn cần cấu trúc & màu.
Thời điểm “tiền ra hoa” – Shikimate tăng Phe trước Trp
Giai đoạn: trước khi nụ bung – 7 đến 14 ngày
Cần gì?
-
Cây phải tăng áp suất turgor, cứng mô, vận chuyển dinh dưỡng mạnh.
-
Hoa chuẩn bị hình thành sắc tố, lignin, flavonoid bảo vệ nụ.
Do đó:
-
Shikimate ưu tiên nhánh Phe, vì:
-
Phe → Cinnamic acid → Coumaroyl-CoA → Flavonoid: chống tia UV, làm dày lớp biểu bì nụ.
-
Phe → SA (Salicylic acid): kích hoạt miễn dịch nội sinh chống nấm–vi khuẩn ở nụ non.
-
Phe → Lignin nhẹ trong cuống hoa → chống rụng cơ học.
-
=> Phe = “hạ tầng” cho hoa.
Nếu cây thiếu Phe ở giai đoạn này → hoa yếu, cuống mềm, nụ rụng, nấm dễ xâm nhập.
📍Bón MKP + Mg + Fe + amino tổng hợp (chưa Trp) → giúp Shikimate “nuôi Phe”.
–
Khi hoa bắt đầu nở – Shikimate “rẽ” sang Trp
Giai đoạn: ra hoa – thụ phấn – đậu trái
Lúc này:
-
Cây tăng mạnh hormone IAA (auxin) để kéo dinh dưỡng đến bầu hoa,
-
Kích hoạt enzyme TAA1/YUC, mở nhánh Trp → IAA.
-
Đồng thời, Trp còn đi vào:
-
Serotonin – Melatonin: cân bằng nhịp sáng–tối, ổn định năng lượng quang hợp.
-
Indole alkaloid: chống stress do thiếu nước, ánh sáng, hoặc phấn hoa khô.
-
→ Trp trở thành nhánh chủ lực của Shikimate trong pha ra hoa.
–
Khi hoa → quả non: Phe “trở lại”, nhưng với vai trò chống rụng & bảo vệ
Khi đậu trái, auxin (IAA) giảm dần, nhưng cây bắt đầu phải “đóng khung” mô quả.
-
Shikimate lại “chia dòng”:
-
Phe → lignin + flavonoid → cứng vỏ, kháng nấm.
-
Trp → auxin duy trì cân bằng dinh dưỡng giữa hạt–vỏ–cuống.
-
Nói cách khác: Phe giữ quả, Trp giữ tín hiệu.
–
Tóm tắt logic Shikimate theo giai đoạn sinh lý
| Giai đoạn | Mục tiêu sinh học | Nhánh Shikimate ưu tiên | Dấu hiệu ngoài đồng | Ứng dụng |
|---|---|---|---|---|
| Sinh trưởng dinh dưỡng | Xây mô, chống stress | Tyr > Phe | Lá xanh, dày, ít cháy nắng | Mg, Mn, Zn, Fe, amino tổng hợp |
| Tiền ra hoa | Cứng mô, tăng miễn dịch | Phe | Cuống cứng, nụ khỏe, ít rụng | MKP + Mg + HaifaStim Force |
| Ra hoa – thụ phấn | Tín hiệu hormone, định hoa | Trp ↑↑, Phe ↓ nhẹ | Hoa nở đều, phấn sống tốt, bầu no | HaifaStim Energy (Trp + amino) + Ca + B |
| Đậu trái non | Bảo vệ mô, dẫn dinh dưỡng | Phe & Trp song hành | Cuống không đen, vỏ bóng | SOP + Ca(NO₃)₂ + amino |
| Nuôi trái chín | Màu – vị – chất lượng | Phe mạnh lại | Trái lên màu, vị đậm, bền trái | MKP + K₂SO₄ + humic/fulvic |
–
Vì sao Shikimate “biết” chuyển nhánh?
Shikimate không có “ý chí”, nhưng enzyme đầu nguồn (chorismate mutase & aminotransferase) được điều hòa bởi tín hiệu hormone & năng lượng:
| Tín hiệu | Tác động lên Shikimate |
|---|---|
| Auxin (IAA) | Kích hoạt nhánh Trp (tự khuếch đại) |
| Cytokinin | Ức chế nhánh Phe, giữ năng lượng cho sinh sản |
| SA (salicylic acid) | Kích hoạt nhánh Phe khi bị tấn công |
| ROS (ánh sáng mạnh, stress) | Làm “chệch dòng” sang Phe để tăng flavonoid |
| Sucrose & NADPH | Nếu dồi dào → Trp ưu tiên; thiếu → Phe ưu tiên |
👉 Vì vậy, việc bón sai chất hoặc thời điểm (ví dụ cho amino Trp khi cây chưa đủ năng lượng)
→ khiến Shikimate bị “phân vân” → hoa không đều, rụng nụ.
–
Kết luận “ngắn mà nhớ mãi”
| Câu khóa | Ý nghĩa |
|---|---|
| “Phe xây hoa – Trp mở hoa.” | Phe tạo khung – Trp truyền tín hiệu. |
| “Thiếu Phe: hoa yếu – Thiếu Trp: hoa câm.” | Một bên khung, một bên hormone. |
| “Shikimate không chọn theo ta – nó chọn theo năng lượng.” | Năng lượng quyết định nhánh hoạt động. |
–
Gợi ý ứng dụng thực hành
Chương trình 10 ngày trước & trong nở hoa:
-
Ngày 1–3: MKP 0.3% + HaifaStim Force → tăng Phe.
-
Ngày 4–6: Ca(NO₃)₂ + B + HaifaStim Energy (0.2%) → bật Trp.
-
Ngày 7–10: SOP + humic nhẹ → giữ Phe & Trp cân bằng.
→ Hoa khỏe, phấn mạnh, bầu no, tỷ lệ đậu cao, không rụng non.
–
3. Một loại thuốc nông dược có thể làm tắc Shikimate không?
Đây chính là chìa khóa để phân biệt nông nghiệp sinh học và nông nghiệp hóa học.
Nếu hiểu được nó, ta sẽ biết vì sao cây “chết không cần cháy”, vì sao lá “vẫn xanh mà Shikimate bị tắt,” và vì sao nhiều loại thuốc, kể cả thuốc cỏ, thuốc nấm, thuốc sâu đều ảnh hưởng đến trí tuệ sinh học của cây.
Giờ ta cùng đi sâu, từng tầng — sinh hóa, cơ chế, phản ứng – và hậu quả trong đất và cây.
Một số nhóm thuốc nông dược (đặc biệt là thuốc cỏ Glyphosate, thuốc nấm nhóm Triazole, và một số thuốc sâu ức chế enzyme P450 hoặc nitrate reductase) có thể làm tắc hoặc chậm con đường Shikimate.
–
Glyphosate – “dao chém giữa đường Shikimate”
🔹 Cơ chế:
Glyphosate (tên hóa học: N-phosphonomethylglycine) là thuốc cỏ toàn diện,
và đích tác động chính của nó chính là Shikimate pathway, cụ thể:
-
Nó ức chế enzyme EPSPS (5-enolpyruvylshikimate-3-phosphate synthase).
-
Enzyme này chịu trách nhiệm nối shikimate-3-phosphate (S3P) và phosphoenolpyruvate (PEP) → tạo 5-enolpyruvylshikimate-3-phosphate (EPSP).
-
Khi EPSPS bị ức chế:
-
Chuỗi dừng lại, không tạo được chorismate → cây mất Phe, Tyr, Trp.
-
Không còn flavonoid, lignin, auxin, SA.
-
Cây vẫn xanh 3–5 ngày (vì còn dự trữ), sau đó chết “trí tuệ” rồi chết thể xác.
-
🔹 Hậu quả sinh lý:
| Biểu hiện | Giải thích |
|---|---|
| Lá vẫn xanh 3–5 ngày | Chlorophyll chưa phân hủy |
| Sau đó lá vàng, mô mềm | Mất lignin, tế bào vỡ |
| Cuối cùng cây chết khô | Không còn amino thơm, ROS phá hủy mô |
🔹 Trong đất:
-
Glyphosate bám vào khoáng P, Fe, Mn → “trói” chúng lại → làm đất thiếu dinh dưỡng hữu hiệu.
-
Giết vi sinh EPSPS dương tính, đặc biệt là Bacillus, Pseudomonas, Streptomyces — nhóm vi sinh sản xuất hormone và kháng sinh tự nhiên.
-
Vi sinh yếm khí tăng mạnh → sinh H₂S, CH₄, NH₃ → “đất hôi, cây ngu”.
🔥 Kết quả: Shikimate chết, nhưng vi sinh kỵ khí sống → đất “đang phân hủy sự sống.”
–
Thuốc nấm nhóm Triazole – “làm chậm vòng carbon của Shikimate”
Triazole (ví dụ: Hexaconazole, Propiconazole, Tebuconazole, Difenoconazole)
được thiết kế để ức chế enzyme CYP51 (lanosterol demethylase) trong nấm.
Nhưng trong thực vật, các enzyme CYP450 (P450 oxygenase) tương tự cũng đóng vai trò trong chuyển hóa Shikimate → Phe/Tyr/Trp.
🔹 Cơ chế:
-
P450 bị ức chế → giảm hydroxyl hóa & khử trong các bước cuối của Shikimate → chậm tổng hợp flavonoid & lignin.
-
Kết quả:
-
Lá vẫn xanh → nhưng lá mỏng, ánh sáng xuyên qua mạnh.
-
Cây “sạch nấm” → nhưng giảm đề kháng nội sinh, dễ bị nấm trở lại sau 10–15 ngày.
-
Rễ phát triển chậm, giảm “hơi thở” hiếu khí.
-
🔹 Hậu quả:
-
Giống như Shikimate “bị kìm hãm” nhưng không chết hẳn.
-
Nếu dùng liên tục → cây “câm miễn dịch,” lá nhợt, trái mất hương.
⚠️ Triazole = thuốc làm cây “ngoan” nhưng mất linh hồn.
–
Thuốc sâu nhóm ức chế Cytochrome P450 hoặc nitrate reductase
Ví dụ:
-
Imidacloprid, Thiamethoxam (Neonicotinoid)
-
Chlorfenapyr, Pyridaben, Fenpyroximate, Spiromesifen (ức chế P450)
-
Một số thuốc trừ cỏ chứa ammonium hoặc urea dạng amide
🔹 Cơ chế:
-
P450 là “mạng lưới enzyme khử oxy” trong cây – liên quan đến cả Shikimate, SA, auxin.
-
Khi bị ức chế → cây không thể chuyển hóa Trp thành IAA, hoặc Phe thành SA.
-
Một số ức chế cả nitrate reductase → mất NADPH cân bằng → PEP không tái tạo được.
🔹 Hậu quả:
| Hiện tượng | Nguyên nhân |
|---|---|
| Lá xanh đậm giả, rễ chậm, hoa ít | Nitrate reductase yếu, carbon lệch |
| Lá dày bất thường | P450 bị ức chế, auxin thấp |
| Trái dễ sượng, thiếu hương | Shikimate không vào nhánh Trp |
–
Thuốc diệt cỏ khác – làm tắc Shikimate gián tiếp
| Nhóm | Tên hoạt chất | Ảnh hưởng |
|---|---|---|
| Glufosinate | (PPT, Basta) | Ức chế glutamine synthetase → tích NH₄⁺ → giảm pH rễ, tắc hô hấp, Shikimate chậm |
| Paraquat, Diquat | (Gramoxone) | Tạo ROS quá mạnh → phá diệp lục, oxy hóa NADPH → Shikimate không có năng lượng khử |
| Atrazine, Simazine | (Triazine) | Chặn PSII → không tạo ATP/NADPH → Shikimate “mất điện” |
| 2,4-D, MCPA | (Auxin giả) | Kích Trp–IAA giả → enzyme nội sinh dừng hoạt động thật, “hormone loạn” |
🎯 Điểm chung: hầu hết thuốc cỏ & thuốc nấm đều gián tiếp hoặc trực tiếp ảnh hưởng đến Shikimate, vì nó là đường duy nhất nối carbon – năng lượng – amino thơm.
–
Biểu hiện ngoài đồng – nhận biết Shikimate “bị tắc do thuốc”
| Dấu hiệu thực tế | Cơ chế |
|---|---|
| Lá xanh lạnh, dày giả, không đàn hồi | P450 bị ức chế, lignin – flavonoid thấp |
| Lá chuyển xanh nhợt, rễ vàng nâu | EPSPS tắc, thiếu Phe/Tyr/Trp |
| Hoa ít, phấn yếu | Trp → IAA ngừng, hormone lệch |
| Trái mất mùi, vị nhạt | Phe → phenolic tắc, flavonoid giảm |
| Đất hôi, vi sinh chết | Glyphosate/Triazole giết EPSPS vi sinh |
–
Giải pháp phục hồi sau thuốc làm tắc Shikimate
💧 Ngày 1 (24h sau phun thuốc)
Khử độc & “rửa” enzyme:
-
-
Tưới Ca(NO₃)₂ Agri 0.3% + humic + EM hiếu khí, giúp O₂ tái sinh trong đất, và nitrat “dẫn khí” cho rễ.
-
Bón hữu cơ tơi, không ủ tươi.
-
-
Phá kỵ khí, khử H₂S, CH₄.
→ Mục tiêu: Mở hô hấp để PEP tái tạo được.
Ngày 2-3:
-
-
Phun Mg(NO₃)₂ + Mn + Fe-EDDHA để bật NADPH.
Cấp cứu năng lượng: Phun mannitol/betaine (HaifaStim Force) để tái cân bằng electron.→ Mục tiêu: Cho Shikimate có “điện” để chạy.
-
Ngày 4-5:
Khôi phục amino thơm:
-
-
Sau 3–5 ngày, phun HaifaStim Energy (chứa Trp, Phe tự do).→ cung nguyên liệu cho enzyme EPSPS và PAL hoạt động lại.
-
Giúp enzyme Shikimate “nhớ lại cách hoạt động.”
-
Ngày 6-7:
Ổn định nitrat – P:
-
-
MKP liều nhẹ 0.3% + Ca(NO₃)₂ luân phiên 5–7 ngày.–> để tái tạo PEP–E4P từ glucose.
-
👉 Sau 10–15 ngày, lá sẽ dày thật, rễ trắng, mùi đất tươi, đó là dấu hiệu Shikimate hoạt động trở lại.
–
Chú ý: hạn chế pha chung thuốc BVTV với phân bón
vì bản chất: thuốc = ức chế, phân = kích hoạt
-
Thuốc nông dược (dù là sâu, nấm, cỏ) → ức chế một enzyme hoặc màng tế bào sinh vật.
-
Phân bón → kích hoạt enzyme, tạo năng lượng, nuôi tế bào.
👉 Hai luồng này đi ngược nhau hoàn toàn.
Khi pha chung, enzyme thực vật bị rối: không biết phải chạy hay dừng → Shikimate “tắc nghẽn logic.”
–
Phân tích hóa học: tại sao Shikimate bị ảnh hưởng?
| Thành phần | Vai trò | Khi pha chung thuốc thì… | Hậu quả |
|---|---|---|---|
| PEP / E4P / ATP (nội sinh của cây) | nguồn năng lượng Shikimate | bị chiếm electron bởi gốc Cl, S, P từ thuốc | Shikimate mất điện, dừng phản ứng |
| pH dung dịch | ổn định enzyme EPSPS, PAL | đa số thuốc cần pH <6, còn phân nitrat–phosphate cần pH >6 | enzyme bị biến tính |
| Chelate Fe–Mn–Zn | dẫn truyền electron trong Shikimate | bị phá vỡ bởi gốc Triazole, Dithiocarbamate, hoặc lưu huỳnh | thiếu năng lượng khử |
| Amino tự do (từ phân hữu cơ, HaifaStim) | nguyên liệu cho Shikimate | phản ứng Maillard với thuốc chứa Cu, S, hoặc phosphonate | amino bị “cháy đen”, mất hoạt tính sinh học |
Tức là, khi ta pha chung, thuốc không hỏng, phân không hỏng, nhưng “cây mất khả năng giải mã tín hiệu sinh học.”
Cây vẫn xanh – nhưng Shikimate bị tắc tạm thời 3–5 ngày.
–
Những nhóm tuyệt đối không nên pha chung với phân bón
| Nhóm thuốc | Ví dụ hoạt chất | Vì sao không nên pha |
|---|---|---|
| Thuốc cỏ | Glyphosate, Glufosinate, Paraquat | diệt EPSPS → tắt Shikimate ngay lập tức |
| Thuốc nấm Triazole | Hexaconazole, Propiconazole, Difenoconazole | ức chế P450 → chặn enzyme Shikimate bậc 2 |
| Thuốc đồng & lưu huỳnh | Copper oxychloride, Mancozeb, Sulfur | oxi hóa amino & chelate, phá NADPH |
| Thuốc sâu Neonicotinoid | Imidacloprid, Thiamethoxam | giảm nitrate reductase → PEP tái tạo chậm |
| Phosphonate | Aliette (Fosetyl-Al) | tạo môi trường pH acid → biến tính enzyme PAL, EPSPS |
–
Một số “pha được nhưng phải thông minh”
| Kết hợp | Điều kiện | Cách làm an toàn |
|---|---|---|
| Thuốc sinh học (Bacillus, nấm đối kháng) + Amino | pH 6.0–7.0 | OK, nếu không có đồng/lưu huỳnh |
| Thuốc sâu sinh học (Abamectin, Emamectin) + MKP hoặc Ca(NO₃)₂ | pH trung tính, không dùng nước cứng | OK, phun buổi sáng, không nắng gắt |
| Phân bón lá + thuốc nấm hóa học | Chỉ khi cần “giữ lá sống” trong 1–2 ngày | Pha loãng 1/2 liều, kiểm tra pH <6.5, thêm 1% đường mía hoặc mannitol để giảm stress |
👉 Nói cách khác: được pha = tạm thời an toàn về hóa học, nhưng không an toàn về sinh học.
Shikimate vẫn bị “đè” nhẹ trong 1–2 ngày.
“Đừng pha chung – hãy pha cách thời gian 24–48h, để Shikimate có cơ hội thở.” Nếu bạn vẫn muốn pha hãy biết rõ cơ chế hóa học của từng hoạt chất.
–
Shikimate là ngọn đèn của sự sống.
Một số thuốc “diệt cỏ” hay “diệt bệnh” không giết cây ngay,
nhưng thổi tắt ngọn đèn đó – để cây xanh mà ngu, sống mà không biết sống.
Vì vậy, nông nghiệp sinh học không chỉ là bỏ thuốc,
mà là khôi phục trí tuệ của đất và cây, để Shikimate hát lại —
nơi carbon biến thành hương, màu, và ý chí sinh tồn.
–
4. Shikimate trong vi sinh vật đất có liên kết năng lượng với Shikimate của cây không?
Nếu ta hiểu được mối liên kết Shikimate giữa cây và vi sinh vật đất, thì ta hiểu “sợi dây thần kinh sinh học” nối rễ – đất – trời.
Nó là nền tảng thật sự của nông nghiệp sinh thái, nơi cây không sống một mình, mà hít thở – suy nghĩ – trao đổi năng lượng chung với đất.
Giờ ta cùng “đi xuyên qua đất” – giải mã từng tầng:
4.1 Shikimate – con đường chung của cả sự sống ngoại trừ động vật
-
Gần như mọi sinh vật quang hợp và vi sinh vật đất (vi khuẩn, nấm, tảo, rêu, kể cả cỏ dại) đều có con đường Shikimate.
-
Động vật (và con người) không có Shikimate, nên phải ăn amino thơm (Phe, Tyr, Trp) từ thực vật hoặc vi sinh vật.
👉 Vậy nên, Shikimate chính là “ngôn ngữ trao đổi năng lượng và phòng vệ” giữa cây và vi sinh vật.
Cây và vi sinh vật nói chuyện với nhau bằng chính những sản phẩm từ Shikimate.
–
4.2 Ba điểm “chạm năng lượng” giữa Shikimate của cây và vi sinh vật
4.2.1. Ở vùng rễ – “điểm giao PEP & E4P”
-
Cả cây và vi sinh vật đều dùng PEP (phosphoenolpyruvate) và E4P (erythrose-4-phosphate) làm nguyên liệu đầu vào cho Shikimate.
-
Khi rễ tiết ra đường – amino – acid hữu cơ, vi sinh vật hút lấy carbon này, đồng thời tiết lại enzyme EPSPS, chorismate mutase, tryptophan synthase ra vùng rễ.
-
Những enzyme đó giúp cây chuyển hóa nhanh hơn – tức là Shikimate của vi sinh vật “nối mạch điện” cho cây.
🔋 Nói cách khác: vi sinh vật cho cây mượn điện và enzyme để Shikimate của cây không bị đứt khi stress.
–
4.2. 2. Ở pha chorismate – “ngã ba thông tin”
-
Chorismate là điểm giữa, nơi chia 3 nhánh: Phe, Tyr, Trp.
-
Vi sinh vật đất tạo chorismate ngoại bào (đặc biệt là Pseudomonas, Bacillus, Streptomyces).
-
Cây hấp thu chorismate hoặc SA (salicylic acid) do vi sinh tạo ra → kích hoạt kháng bệnh, dày lá, thơm quả.
🧠 Vi sinh vật “cấy trí nhớ miễn dịch” vào cây thông qua chorismate → SA → hệ thống phòng vệ cảm ứng (SAR – Systemic Acquired Resistance).
–
4.2.3. Ở pha sản phẩm phụ – “trao đổi hormone & tín hiệu”
-
Vi sinh vật tạo ra:
-
Indole acetic acid (IAA) từ Trp → kích rễ cây.
-
Melatonin & phenolic → chống oxy hóa, giúp cây chịu stress.
-
Phylloquinone (vitamin K₁) → xúc tác quang hợp của cây.
-
Siderophore → lấy Fe cho cả hai bên dùng.
-
-
Cây đáp lại bằng:
-
Exudate chứa flavonoid, phenolic → làm tín hiệu cho vi sinh bám rễ.
-
Carbon hữu cơ (đường, acid amin) → “nuôi” lại vi sinh đó.
-
→ Hai bên nuôi và dạy nhau – đúng nghĩa cộng sinh năng lượng sinh học.
–
4.3. Liên kết năng lượng: “hô hấp cộng sinh” giữa Shikimate cây – đất
Tầng sâu sinh hóa:
-
Khi vi sinh vật oxy hóa carbon → tạo NADPH và ATP ngoại bào.
-
Rễ hấp thu tín hiệu NADPH này qua chất tiết flavonoid, nhờ membrane redox system (MRS).
-
MRS là “ổ cắm điện” giữa Shikimate vi sinh và Shikimate cây:
-
vi sinh “cắm điện” vào rễ,
-
cây “sạc lại” năng lượng khử cho Shikimate của chính mình.
-
⚡️ Giống như hệ thống smart grid – vi sinh tạo điện, cây tiêu điện, nhưng khi cây quang hợp mạnh, nó lại “nạp điện ngược” vào vi sinh (trao carbon).
–
4.4 Khi cây stress – vi sinh trở thành “lá phổi của Shikimate”
-
Khi đất thiếu oxy, cây mất hô hấp → NADPH lệch → Shikimate ngừng.
-
Vi sinh vật hiếu khí (Bacillus, Azotobacter, Rhizobium…) lúc này “thở thay cho cây” bằng chuỗi electron hiếu khí.
-
Chúng tái sinh PEP và NADPH quanh rễ → Shikimate cây “bắt tín hiệu” và chạy lại.
👉 Đó là lý do khi tắt kỵ khí – bật rễ trắng, Shikimate cây “thức dậy nhanh” chỉ sau 3–4 giờ.
Vì vi sinh đang “đưa oxy – điện – chorismate” cho cây.
–
4.5 Khi Shikimate cây bị thuốc tắt – vi sinh cũng “nghẹt điện”
-
Thuốc như glyphosate không chỉ ức chế EPSPS trong cây, mà còn diệt luôn vi sinh có con đường này.
-
Khi vi sinh chết → chorismate trong đất biến mất → SA nội sinh cây sụp → cây mất kháng.
-
Lúc đó, đất hôi (H₂S, CH₄), cây thối rễ, lá “ngu” – biểu hiện điển hình của mất liên kết Shikimate kép.
🌪️ “Cây chết không phải vì độc tố, mà vì mất cộng sinh Shikimate.”
4.6 Cách phục hồi “cộng sinh Shikimate cây – đất”
| Giai đoạn | Hành động | Mục tiêu |
|---|---|---|
| Ngày 1 | Xới thoáng, tưới EM hiếu khí + Ca(NO₃)₂ 0,3% | Mở dòng O₂, tái kích PEP vi sinh |
| Ngày 2–3 | Bón humic/fulvic + Mg–Fe + HaifaStim Force | Bật NADPH cho vi sinh + rễ |
| Ngày 4–5 | Phun HaifaStim Energy (amino + Trp + Phe) | Cung nguyên liệu Shikimate cả 2 hệ |
| Ngày 6–7 | Giữ ẩm ổn định, ánh sáng tốt | Ổn định chuỗi hô hấp cộng sinh |
→ Sau 7 ngày, Shikimate cây và vi sinh “nối mạch điện trở lại.”
Lá dày, thơm, rễ trắng, đất có mùi ngọt — đó là dấu hiệu liên kết năng lượng đã hồi.
–
“Shikimate không chỉ là con đường hóa học, mà là mạch sống thống nhất giữa cây và đất.”
Cây nghĩ – vi sinh thở – đất ghi nhớ.
Khi một bên tắt, cả hệ thống quên cách sống.
Khi cả hai bật lại, tự nhiên trở về trạng thái có trí tuệ.
Seiko chúc bạn hiểu và cùng Seiko chăm tốt vườn của mình một cách bền vững.
Bạn có bất kỳ câu hỏi nào, liên hệ về Seiko nhé. Cùng nhau chúng ta xây dựng nông nghiệp tốt hơn, bền vững hơn cho hôm nay và mai sau.
Nguồn: Seiko tổng hợp




